|
IZHIKEVICH MODEL FOR ACTION
POTENTIALS AND SPIKE TRAINS |
||
|
ns_Izh002.m
Computation of the firing patterns of a single neuron using the Izhikevich Model |
||
|
This article and the Scripts
are based upon the papers by E. M. Izhikevich Which Model to Use for Cortical Spiking Neurons? IEEE Transactions on Neural Networks.
Vol. 15. No. 5. September 2004. Srdjan Ostojic. Two
types of asynchronous activity in networks of excitatory and inhibitory
spiking neurons. Nature
Neuroscience
Vol. 17. No. April 2014 |
||
|
Most
neurons are excitable in that they can fire a voltage spike when stimulated.
In developing useful brain models we want to account for and explain the
patterns produced by spiking neurons. To do this we must satisfy two
requirements: 1)
The model
must be computationally simple. 2)
The model
must be able to produce the rich firing patterns exhibited by real biological
neurons. The
Hodgkin–Huxley-type models are computationally prohibitive, since they
can be used only to simulate a handful of neurons in real time. However, the
use of integrate-and-fire models are computationally effective, but the
models are unrealistically simple and incapable of producing the rich spiking
and bursting dynamics exhibited by cortical neurons. An
interesting model to consider was proposed by E. M. Izhikevich.
Although this model is not biophysically meaningful, it can be used to
compute a wide range of neuron spiking patterns for cortical neurons. The Izhikevich model presented is biologically plausible as
the Hodgkin–Huxley model, yet as computationally efficient as the
integrate-and-fire model. The value of four parameters a, b,
c and d
used in the model, determines the
spiking and bursting behaviour of the known types of cortical neurons. The
time evolution of the membrane potential v
is described in terms of the
differential equations |
||
|
(1)
(2) The after-spike
resetting relationship is (3) where
u is the membrane recovery
variable. |
||
|
The
dimensions and values of the model parameters are v
membrane potential
[mV] t time [ms] dv/dt time rate of change
in membrane potential
[mV.ms-1 or V.s-1] u
recovery variable
[mV] I external
current input to cell (synaptic currents or injected DC-currents) [A] c1 = 0.04 mV‑1.ms-1 c2 = 5 ms-1 c3 = 140 mV.ms-1 c4 = 1 ms-1 c5
= 1 mV.ms-1.A-1
|
||
|
The membrane recovery variable u provides negative feedback to the membrane
potential v
and accounts for the activation of the K+ currents and
inactivation of the Na+ currents. The term Various choices of the parameters a, b,
c and d
result in the various intrinsic
firing patterns that can be computed. a ~ 0.02 ms-1 determines the time
scale of the recovery variable u.
The larger the value of a
the quicker the recovery. b
~ 0.20 [dimensionless] describes the sensitivity of the recovery variable u to the subthreshold fluctuations of
the membrane potential v.
Larger values of b
couple u
and v more strongly resulting in possible
subthreshold oscillations and low-threshold spiking dynamics. c ~ -65 mV gives the after-spike reset value
of the membrane potential v
caused by the fast high-threshold K+ conductances. d ~ 6 mV describes after-spike reset of the
recovery variable u
caused by slow high-threshold Na+
and K+ conductances. A neuron can be stimulated by the
injection of DC current pulse via an electrode and the response of the
membrane potential recorded. When a step-input current is used to stimulate a
neuron, the neuron continues to fire a sequence of spikes called a spike-train.
From such investigations, it is found that neocortical neurons in the
mammalian brain can be classified into several firing patterns. Most of these
recorded firing patterns can be reproduced computationally using the Izhikevich Model. The
following figures show the spike-train patterns for a step input current that
causes the neuron to continually fire. The spike-train pattern depends upon
the values of the four parameters a,
b, c
and d and
the input current I.
The mscript ns_Izh002.m was used to
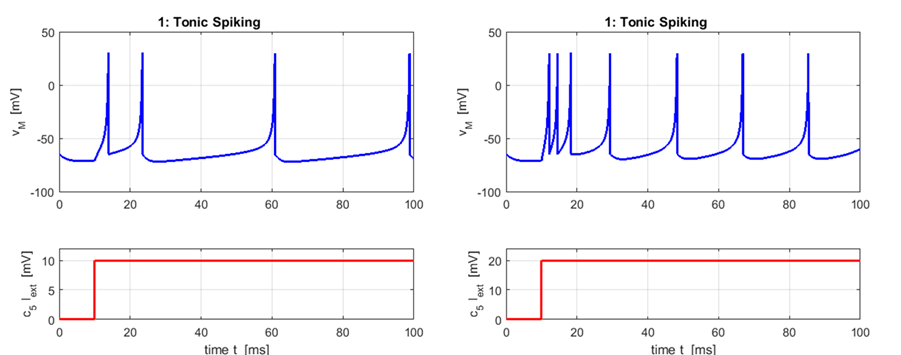
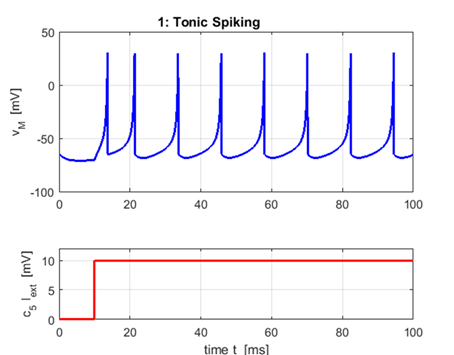
generate the plots. 1. Tonic Spiking and Bursting |
||
|
Excitatory neurons:
Regular
Spiking a = 0.02 b = 0.20 c = -65 d = 6
The most common type of excitatory neurons
in the mammalian neocortex are regular spiking cells that fire spikes with
decreasing frequency. When presented with a prolonged stimulus the neurons
fire a few spikes with short interspike period and
then the period increases. This is called spike frequency adaptation.
The frequency is relatively high at the oneset of
the stimulation and then it adapts. Increasing the strength of the injected DC
current increases the interspike frequency, though
it never becomes too fast because of large spike after hyperpolarizations. |
|
|
|
|
||
|
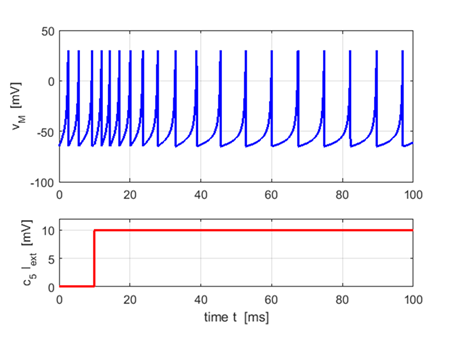
Inhibitory
neurons: Fast Spiking
a = 0.10 (fast recovery) b = 0.20
c = -65 d = 6
Neurons
fire periodic trains of action potentials with extremely high frequency
practically without any adaptation (slowing down) |
||
|
|
||
|
Inhibitory
neurons: Low-Threshold
Spiking a
= 0.02 b
= 0.4 c = -65 d = 2
Neurons
can also fire high-frequency trains of action potentials, but with a
noticeable spike frequency adaptation. These neurons have low firing
thresholds, which is accounted for by b = 0.4 in the model. Because
of large b value hence low threshold, spikes are
produced before external pulse is switched on. |
||
|
|
||
|
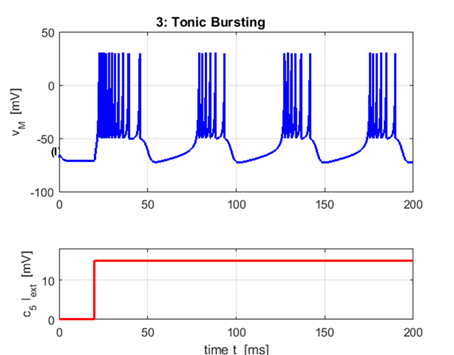
Excitatory
(chattering) neurons: Tonic bursting a = 0.02 b = 0.20
c = -50 (very high reset
voltage) d
= 2 (moderate after-spike jump of u)
Some
excitatory neutrons such as chattering neurons
in a cat cortex fire periodic bursts of closely spaced spikes when
stimulated and may contribute to the gamma frequency oscillations in the
brain. The interburst frequency can be as high as
50 Hz. |
||
|
|
||
|
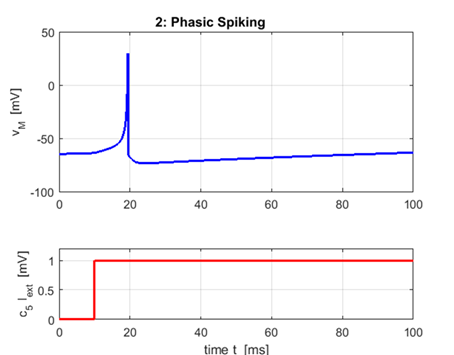
2. Phasic
Spiking and Bursting |
||
|
Phasic
spiking: a
= 0.02 b = 0.25
c = -65 d = 6
A neuron may only fire a single spike at the
onset of the input stimulus and remain quiescent afterwards. Phasic
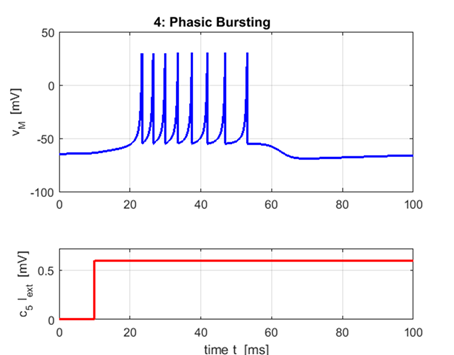
bursters: set of closely spaced spikes are
generated and then remains quiescent afterwards. a = 0.02 b
= 0.25 c = -55 d = 0.05
Phasic bursting importance: · May overcome synaptic failure and reduce
noise. · Postsynpatic signal is stronger than that of a single
spike. · Selective communication between neurons (interspike frequency within the
burst encodes communication channels). |
|
|
|
|
||
|
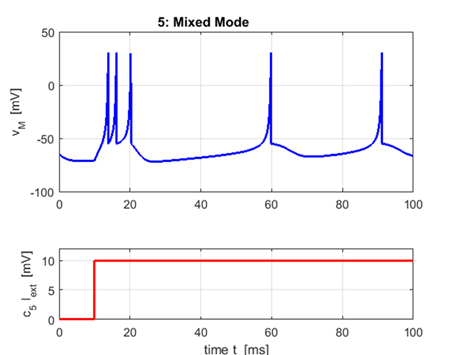
Mixed
Mode a
= 0.02 b = 0.2
c = -55 d = 4
Excitatory neurons in the mammalian cortex
can exhibit a phasic burst at the start of the stimulus and then switch to
the tonic spiking mode. |
||
|
|
||
|
|
||
|
Many more spike-train patterns can be
investigated with different input stimulus profiles using the mscript ns_Izh002.m for the Izhikevich Model. |
||
|
|
||
|
|
||
|
|
||
|
|
||
|
|
||
|
|
||
|
|
||
|
|
||
|
|
||
|
|
||
|
|
||